Patrones de abundancia y diversidad en seis especies Columbidae en el Monte Natural Cupaynicú, Guisa, Cuba
Patterns of abundance and diversity in six Columbidae species in the Cupaynicú Natural Forest, Guisa, Cuba
Elianis Guerra Cañete(1)
Lázara Meilín Infante Dieguez(2)
Sergio Florentino Rodríguez Rodríguez(3)
Idalmis Tamayo Cordovés(4)
Jorge Liusvert Pérez Pérez(5)
Dayana Morejón Rodríguez(6)
(1) Universidad de Granma, Cuba. eguerrac25@gmail.com.
ORCID: https://orcid.org/0009-0006-5028-8403
(2) Universidad de Granma, Cuba. lmeilinid@gmail.com.
ORCID: https://orcid.org/0009-0001-7263-0250
(3) Universidad de Granma, Cuba. sfrodriguez1964@gmail.com
ORCID: https://orcid.org/0000-0003-2923-5092
(4) Universidad de Granma, Cuba. idajorgitin@gmail.com
ORCID: https://orcid.org/0009-0009-4243-9189
(5) Universidad de Granma, Cuba. liusvert2021@gmail.com
ORCID: https://orcid.org/0000-0003-3372-7559
(6) Jardín Botánico Cupaynicú, Granma, Cuba. dayanamorejonrodriguez2@gmail.com
ORCID: https://orcid.org/0000-0002-4994-9976
Contacto: eguerrac25@gmail.com
Artículo recibido: 12/septiembre/2025. Aprobado: 05/diciembre/2025
Resumen
Es importante el estudio de la avifauna para los ecosistemas forestales. El estudio evaluó la estructura de la comunidad de aves en un hábitat específico, revelando una dominancia marcada de Columbina passerina, que representó el 56,77 % de los 227 individuos observados, mientras que otras especies como Streptopelia decaocto mostraron proporciones significativamente menores. La prueba de chi-cuadrado indicó una heterogeneidad significativa en la distribución de especies (χ² = 410,67, p < 0,0001), evidenciando que las proporciones no son homogéneas y sugiriendo que factores ecológicos como preferencias de hábitat, disponibilidad de recursos y presiones antropogénicas influyen en esta distribución desigual. El análisis por pares confirmó diferencias significativas, destacando que cada especie ocupa un nicho ecológico distintivo, salvo algunas con baja potencia estadística por su escasa presencia. Los residuos estandarizados señalaron contribuidores significativos a la desviación global, enfatizando la sobreabundancia de Columbina passerina y la subrepresentación de varias especies raras, posiblemente por competencia, depredación o limitaciones de hábitat. Los índices ecológicos reflejaron una diversidad moderada-baja (Shannon H’=1,19), baja equitatividad (Pielou J’=0,67) y alta dominancia, indicando un ecosistema perturbado o especializado. El análisis de agrupamiento jerárquico distinguió dos grupos claros: uno representando a Columbina passerina como especie dominante y otro agrupando a las especies restantes con bajas proporciones, reforzando la heterogeneidad comunitaria. Estos resultados evidencian un estado de desequilibrio ecológico y subrayan la necesidad de investigaciones futuras para determinar los factores bióticos y abióticos que regulan la estructura comunitaria y sus implicaciones para la conservación de la avifauna en hábitats afectados por antropización.
Palabras claves: comunidad de aves; conservación ambiental; diversidad biológica; nicho ecológico
Abstract
It is important to study avifauna for forest ecosystems. The study evaluated the structure of the bird community in a specific habitat, revealing a marked dominance of Columbina passerina, which accounted for 56.77 % of the 227 individuals observed, while other species such as Streptopelia decaocto showed significantly lower proportions. The chi-square test indicated significant heterogeneity in species distribution (χ² = 410.67, p < 0.0001), demonstrating that proportions are not homogeneous and suggesting that ecological factors such as habitat preferences, resource availability, and anthropogenic pressures influence this uneven distribution. Pairwise analysis confirmed significant differences, highlighting that each species occupies a distinct ecological niche, except for some with low statistical power due to scarce presence. Standardized residuals identified significant contributors to the overall deviation, emphasizing the overabundance of Columbina passerina and the underrepresentation of several rare species, possibly due to competition, predation, or habitat limitations. Ecological indices reflected moderate-low diversity (Shannon H’=1.19), low evenness (Pielou J’=0.67), and high dominance, indicating a disturbed or specialized ecosystem. Hierarchical clustering analysis distinguished two clear groups: one representing Columbina passerina as the dominant species and another grouping the remaining species with low proportions, reinforcing the community heterogeneity. These results reveal a state of ecological imbalance and underscore the need for future research to identify biotic and abiotic factors regulating community structure and their implications for the conservation of avifauna in habitats affected by anthropization.
Key words: bird community; environmental conservation; biological diversity; ecological niche
Introducción
La biodiversidad global se encuentra en un momento crítico, con aproximadamente un millón de especies de plantas y animales en peligro de extinción debido a las actividades humanas como la destrucción de hábitats, el cambio climático y la contaminación. Esta pérdida acelerada representa una amenaza directa para la salud de los ecosistemas, la seguridad alimentaria y el bienestar humano, por lo que requiere acciones urgentes y coordinadas a nivel mundial para promover un desarrollo sostenible y la conservación ambiental (ONU, 2025).
La utilización de la fauna silvestre, a menudo desordenada y no sostenible, aunada a la creciente pérdida y deterioro de sus hábitats, da lugar a una degradación del recurso en diversidad, abundancia y distribución, lo cual demanda una enérgica acción por parte del estado (Mancina y Cruz, 2017).
Las áreas protegidas son las herramientas más ensayadas para la conservación de la biodiversidad, aunque ésta sigue en declive en todo el mundo. Su concepción original ha evolucionado para incorporar aspectos territoriales, sociales y económicos a los primigenios objetivos de conservación. El cambio global acelera la necesidad de abordar de forma más holística e innovadora su planificación y gestión. La transferencia del conocimiento científico a la toma de decisiones y la implicación de la sociedad son cruciales (Mugica et al., 2019).
Dentro de los vertebrados terrestres, las aves representan la clase más diversa, con cerca de 11 000 especies a nivel global (Billerman et al., 2022). Las aves brindan importantes servicios ecosistémicos y son empleadas como bioindicadores por su fácil identificación y su posición dentro de las redes tróficas de las comunidades (González et al., 2017). Debido a que las aves han experimentado un declive poblacional global en las últimas cuatro décadas, lo que afecta negativamente su conservación, se requiere dilucidar las amenazas y desarrollar acciones de conservación eficaces (Rosenberg et al., 2019).
En el caso de la avifauna cubana, existen amplios vacíos de conocimiento, no solo sobre aves migratorias, sino también sobre las especies residentes y endémicas. Aunque existen trabajos que abordan aspectos de la ecología de endémicos cubanos, estos aún son escasos. Este hecho resalta la importancia de mantener esfuerzos de monitoreo de las poblaciones de nuestras aves a largo plazo, que permitan diseñar estrategias de conservación bien definidas (Arañaburo, 2024).
Además en el entorno del Refugio de la Vida Silvestre Monte Natural Cupaynicú en Guisa, provincia Granma, Cuba, no están claramente establecidos los patrones de composición y diversidad de estas comunidades en el área estudiada.
En tal sentido, este trabajo tuvo como objetivo evaluar los patrones de abundancia y diversidad en seis especies de la familia Columbidae en el Monte Natural Cupaynicú, Guisa, Cuba.
Materiales y métodos
El estudio se realizó en el Refugio de la Vida Silvestre Monte Natural Cupaynicú, con una extensión de 54,58 ha, localizado en los 20°16´54.1” y 20°16´23.9” de latitud norte y los 76°34´8.9” y los 76°34´58.7” de longitud oeste, en la vertiente norte de las pre montañas de la Sierra Maestra en Guisa, Granma, Cuba.
Se realizó un análisis de proporciones múltiples para evaluar la estructura de la comunidad de aves en un hábitat determinado, utilizando datos de conteo de seis especies: Patagioenas squamosa (42), Patagioenas leucocephala (9), Streptopelia decaocto (2), Columbina passerina (130), Geotrygon montana (41) y Zenaida macroura (5).
Inicialmente, se calcularon las proporciones de cada especie respecto al total de individuos observados (n = 227) y sus intervalos de confianza del 95 % mediante el método exacto de Clopper-Pearson. Posteriormente, se realizó una prueba de chi-cuadrado de bondad de ajuste para evaluar la hipótesis nula de homogeneidad de proporciones entre todas las especies, comparando las frecuencias observadas con las esperadas bajo una distribución equitativa.
Para las comparaciones por pares entre especies, se empleó la prueba exacta de Fisher debido a la presencia de conteos bajos en algunas especies. Se construyeron tablas de contingencia 2×2 para cada par de especies, evaluando la diferencia en sus proporciones. Los valores p resultantes se ajustaron mediante el método de Benjamini-Hochberg (FDR) para controlar la tasa de falsos descubrimientos por comparaciones múltiples.
Se realizó un análisis de residuos estandarizados para identificar las especies que contribuían significativamente a la chi-cuadrado global, considerando como contribución relevante aquellos residuos con valor absoluto superior a 1,96. Adicionalmente, se calcularon varios índices de diversidad ecológica: el índice de Shannon (H') para evaluar la diversidad considerando riqueza y equitatividad; el índice de Simpson (1-D) para medir la dominancia de especies; la equitatividad de Pielou (J') para cuantificar la distribución de individuos entre especies; la riqueza específica (S) como número total de especies; y la dominancia (D) como la proporción máxima observada.
Para identificar patrones de similitud entre especies, se realizó un análisis de agrupamiento jerárquico utilizando el método de enlace completo y la distancia euclidiana entre las proporciones de las especies. El umbral de corte fue al 50,0 % de la distancia máxima euclidiana.
El análisis se implementó en el entorno estadístico R 4.5.1, utilizando los paquetes binom para el cálculo de intervalos de confianza mediante el método exacto de Clopper-Pearson; FSA y Vegan para el cálculo de índices de diversidad (Shannon, Simpson) y equitatividad de Pielou. Se empleó la prueba exacta de Fisher para las comparaciones de proporciones por pares entre especies.
Para el cálculo de los residuos estandarizados o residuos de Pearson no se utilizó un paquete específico de R, se implementó directamente mediante operaciones aritméticas básicas en R utilizando las funciones base del lenguaje. El método implementado corresponde al procedimiento estándar para obtener residuos estandarizados en tablas de contingencia, que es fundamental para identificar las celdas que contribuyen significativamente al estadístico Chi-cuadrado global, como:
Residuo estandarizado = (Valor observado - Valor esperado) / √(Valor esperado).
El paquete utilizado para el análisis de agrupamiento (cluster) en el script fue Cluster, que es un paquete especializado de R para análisis de agrupamiento y clasificación. Este paquete proporciona funciones avanzadas para métodos de clustering jerárquico y particional, aunque en el análisis específico se utilizó principalmente la función hclust() del paquete base stats (que viene preinstalado con R) para realizar el agrupamiento jerárquico mediante el método de enlace completo. Para el dendrograma se utilizó ggdendro.
Resultados y discusión
La prueba global de Chi-cuadrado reveló una desviación significativa de la homogeneidad esperada bajo una distribución uniforme (p = 0,0001), indicando que las proporciones no son iguales entre las especies (Tabla 1), rechazando la hipótesis nula de distribución equitativa. Esta heterogeneidad sugiere que factores ecológicos, como preferencias de hábitat, disponibilidad de recursos o presiones antropogénicas, podrían influir en la abundancia diferencial observada. Por ejemplo, especies como C. passerina muestran una alta adaptabilidad a hábitats abiertos y alterados por humanos, lo que podría explicar su dominancia en entornos perturbados
El análisis de las proporciones de especies de aves en el hábitat estudiado reveló una distribución marcadamente heterogénea. Columbina passerina fue la especie dominante, representando el 56,77 % del total de individuos observados (n = 227), con un intervalo de confianza del 95 % entre 50,08 % y 6,28 %. En contraste, S. decaocto mostró la proporción más baja (0,87 %, IC 95 %: 0,11 % -3,12 %).
| Especie | Conteo | Proporción | Ic 95 % |
|---|---|---|---|
| Patagioenas squamosa | 42 | 0,1834 | (0,1355, 0,2397) |
| Patagioenas leucocephala | 9 | 0,0393 | (0,0181, 0,0733) |
| Streptopelia decaoto | 2 | 0,0087 | (0,0011, 0,0312) |
| Columbina passerina | 130 | 0,5677 | (0,5008, 0,6328) |
| Geotrygon montana | 41 | 0,1790 | (0,1317, 0,2349) |
| Zenaida macroura | 5 | 0,0218 | (0,0071, 0,0502) |
| Significación global | NA | NA | p-valor: 0,0001 |
Tabla 1. Proporciones de especies de aves e intervalos de confianza del 95 % (método Clopper-Pearson)**
La marcada diferencia en abundancia entre Columbina passerina y las demás especies podría explicarse por varios factores ecológicos. En primer lugar, es posible que esta especie presente adaptaciones morfológicas, fisiológicas o comportamentales que le permiten explotar más eficientemente los recursos disponibles en el hábitat estudiado. Estudios previos han documentado que Columbina passerina es una especie altamente adaptable a ambientes modificados por humanos, lo que podría explicar su éxito en este hábitat particular.
Para profundizar en las diferencias específicas, se realizaron comparaciones por pares utilizando la prueba exacta de Fisher, con ajuste por comparaciones múltiples mediante el método de falsa tasa de descubrimiento (FDR). La Tabla 2 detalla estos resultados, destacando pares con diferencias significativas (FDR<0,05), como Columbina passerina que difirió significativamente de todas las demás especies excepto de Stretopelia decaocto (FDR = 0,4645), lo que podría indicar similitudes ecológicas limitadas.
Pares como Patagioenas squamosa vs. Geotrygon montana también mostraron diferencias marcadas (FDR = 0,0010), sugiriendo nichos diferenciados; Geotrygon montana prefiere bosques tropicales y es sensible a la fragmentación, limitando su abundancia en áreas alteradas. Sin embargo, varios pares con conteos bajos, como Patagioenas leucocephala vs. Zenaida macroura (FDR = 1,0000), no exhibieron diferencias, posiblemente debido a la baja potencia estadística en muestras pequeñas.
| Comparación entre pares de especies | Valor p (No ajustado) |
Valor p (FDR) |
Significativo (FDR < 0,05) |
|---|---|---|---|
| Patagioenas squamosa vs Patagioenas leucocephala | 0,3716 | 0,6968 | No |
| Patagioenas squamosa vs Streptopelia decaoto | 1,0000 | 1,0000 | No |
| Patagioenas squamosa vs Columbina passerina | 0,0000 | 0,0000 | Sí |
| Patagioenas squamosa vs Geotrygon montana | 0,0002 | 0,0010 | Sí |
| Patagioenas squamosa vs Zenaida macroura | 0,5875 | 0,8828 | No |
| Patagioenas leucocephala vs Streptopelia decaoto | 1,0000 | 1,0000 | No |
| Patagioenas leucocephala vs Columbina passerina | 0,0004 | 0,0016 | Sí |
| Patagioenas leucocephala vs Geotrygon montana | 0,3684 | 0,6968 | No |
| Patagioenas leucocephala vs Zenaida macroura | 1,0000 | 1,0000 | No |
| Streptopelia decaoto vs Columbina passerina | 0,1858 | 0,4645 | No |
| Streptopelia decaoto vs Geotrygon montana | 1,0000 | 1,0000 | No |
| Streptopelia decaoto vs Zenaida macroura | 1,0000 | 1,0000 | No |
| Columbina passerina vs Geotrygon montana | 0,0000 | 0,0000 | Sí |
| Columbina passerina vs Zenaida macroura | 0,0142 | 0,0427 | Sí |
| Geotrygon montana vs Zenaida macroura | 0,5885 | 0,8828 | No |
Tabla 2. Comparaciones por pares entre especies (prueba exacta de Fisher con ajuste FDR)**
Estos hallazgos subrayan la necesidad de muestreos adicionales para confirmar patrones en especies raras, como Patagioenas leucocephala, que depende de manglares para anidar y bosques para alimentarse, enfrentando amenazas por pérdida de hábitat y caza.
El análisis de residuos estandarizados complementa estas comparaciones, identificando contribuciones individuales a la desviación global de la Chi-cuadrado. La Tabla 3 presenta estos residuos, donde valores absolutos superiores a 1,96 indican contribuciones significativas (aproximadamente equivalente a p< 0,05 en distribución normal).
La especie Columbina passerina mostró el residuo positivo más alto (14,86), confirmando su sobreabundancia relativa, mientras que Streptopelia decaocto, Zenaida macroura y Patagioenas leucocephala exhibieron residuos negativos, sugiriendo subabundancia. En tanto, P. squamosa y G. montana presentaron residuos no significativos, indicando que sus proporciones se alinean más con lo esperado bajo homogeneidad. Mientras que la especie P. squamosa es común en bosques caribeños, pero su abundancia puede verse afectada por la caza y fragmentación (Tabla 3).
| Patagioenas squamosa | Patagioenas leucocephala | Streptopelia decaocto | Columbina passerina | Geotrygon montana | Zenaida macroura |
|---|---|---|---|---|---|
| 0,62 | -4,72 | -5,85 | 14,86 | 0,46 | -5,37 |
Especies con contribuciones significativas (residuo >1,96): P. leucocephala, S. decaocto, C. passerina y Z. macroura.
Tabla 3. Residuos estandarizados por especie (redondeados a dos decimales).
La ausencia de pares de especies con proporciones estadísticamente similares después del ajuste por comparaciones múltiples sugiere que cada especie ocupa un nicho ecológico distintivo en la comunidad, con excepción de las especies menos abundantes que podrían compartir recursos o estrategias ecológicas similares. Sin embargo, el bajo poder estadístico debido a los reducidos conteos de algunas especies (Streptopelia decaocto, Zenaida macroura) limita la capacidad de detectar diferencias significativas entre estas especies raras.
Los residuos estandarizados negativos observados en Patagioenas leucocephala, Streptopelia decaocto y Zenaida macroura indican que estas especies están menos representadas de lo esperado bajo una distribución equitativa. Esto podría atribuirse a factores como presión depredadora, competencia interespecífica, disponibilidad limitada de recursos específicos o requerimientos de hábitat restringidos. Por otro lado, el residuo positivamente significativo de Columbina passerina refuerza su condición de especie dominante en la comunidad.
Análisis de Diversidad de Especies:
Índice de Shannon (H'): 1,1926
Índice de Simpson (1-D): 0,6099
Equitatividad de Pielou (J'): 0,6656
Riqueza específica (S): 6
Dominancia (D): 0,5677 (Columbina passerina)
En términos de diversidad, el índice de Shannon (H' = 1,1926) indica una diversidad moderada-baja, mientras que el de Simpson (1-D = 0,6099) y la equitatividad de Pielou (J' = 0,6656) sugieren una distribución desigual, dominada por pocas especies. La riqueza específica (S = 6) es limitada, y la dominancia (D = 0,5677) recae en Columbina passerina, lo que podría reflejar un ecosistema perturbado o especializado, posiblemente influido por la proximidad a áreas humanas que favorecen especies generalistas. Estos índices apuntan a una comunidad con baja equitatividad, posiblemente influida por factores como la urbanización o la disponibilidad de alimento, que benefician a especies como Streptopelia decaocto, aunque aquí es rara pese a su adaptabilidad a hábitats abiertos y agrícolas.
La baja diversidad observada (H' = 1,1926) y la alta dominancia (D = 0,5677) son características típicas de comunidades en etapas sucesionales tempranas o en ambientes con disturbios frecuentes. Estos patrones también pueden observarse en hábitats fragmentados o con recursos limitados, donde unas pocas especies generalistas dominan la comunidad. Futuros estudios deberían investigar los factores ambientales y bióticos que determinan esta estructura comunitaria, así como evaluar las implicaciones de esta dominancia para la dinámica poblacional a largo plazo y la conservación de las especies menos abundantes.
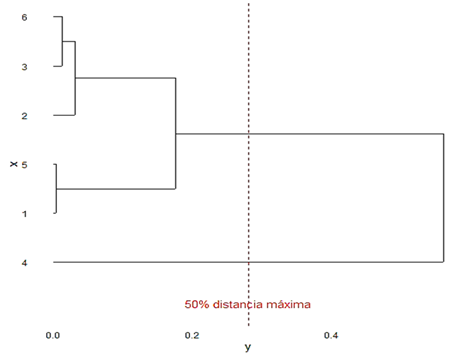
El análisis de agrupamiento jerárquico basado en distancias euclidianas de proporciones, reveló patrones de similitud. El dendrograma con un corte al 50 % de la distancia máxima (0,2795), identificó dos grupos: Grupo 1 (Patagioenas squamosa, Patagioenas leucocephala, Streptopelia decaocto, Geotrygon montana, Zenaida macroura) con proporciones bajas y similares; Grupo 2 (Columbina passerina) como outlier dominante (Figura 1).
Este agrupoamiento sugiere que las especies del Grupo 1 comparten nichos ecológicos o limitaciones similares, como preferencias por bosques (e.g., Geotrygon montana, en bosques tropicales o Patagioenas leucocephala en manglares), mientras que Columbina passerina podría beneficiarse de adaptaciones únicas a hábitats abiertos. No se identificaron pares con similitudes estadísticas (FDR > 0,05) en el análisis por pares, lo que refuerza la heterogeneidad general.
Figura 1. Dendrograma de similitud entre especies en base a sus proporciones por encadenamiento promedio jerárquico y distancia euclidiana
El análisis de agrupamiento confirmó la existencia de dos grupos claramente diferenciados: uno compuesto por la especie dominante y otro que agrupa a todas las especies con abundancias menores. Esta estructura es consistente con lo observado en otras comunidades de aves donde una o pocas especies dominantes ejercen un fuerte efecto competitivo sobre las demás, limitando su abundancia.
Los resultados de este estudio revelan una estructura de comunidad de aves fuertemente dominada por Columbina passerina, que representa más de la mitad de todos los individuos observados. Esta distribución desigual se refleja en los bajos valores de los índices de diversidad y equitatividad, sugiriendo que la comunidad se encuentra en un estado de desequilibrio ecológico o que existen factores ambientales que favorecen selectivamente a esta especie.
Conclusiones
Este estudio revela una comunidad de aves estructurada en torno a una especie dominante, con baja diversidad y alta dominancia. Estos resultados proporcionan una línea base para futuras investigaciones sobre los factores ecológicos que determinan esta estructura y sus implicaciones para la conservación de la avifauna en el hábitat estudiado.
Referencias bibliográficas
Arañburo, L. (2024). Caracterización del ensamble de aves en dos zonas fitogeográficas del Jardín Botánico Nacional de Cuba. Tesis de Diploma, Universidad de La Habana, 57 p.
Billerman, S. M., Keeney, B.K., Rodewald, P.G., & Schulenberg, T.S. (2022). Birds of the world. Cornell Laboratory of Ornithology, Ithaca, NY, USA.
González Rossell, A., Ferro Armas, J., & Rodríguez Cabrera, D. (2017). Aves terrestres. En C. A. Mancina & D. D. Cruz (Eds.), Diversidad biológica de Cuba: métodos de inventario, monitoreo y colecciones biológicas (pp. 413-447). Editorial AMA, La Habana, Cuba.
Mancina, C. A., & Cruz Flores, D. D. (2017). Diversidad biológica de Cuba: métodos de inventario, monitoreo y colecciones biológicas (ISBN 978-959-300-130-4). Editorial AMA, La Habana.
Mugica, S. A., González Rossell, A., Serrano Méndez, H., & Crespo, B. (2017). Las áreas protegidas como herramienta para la conservación de la biodiversidad en Cuba. En Agencia de Medio Ambiente de Cuba (Ed.), Tabloide Áreas Protegidas de Cuba y la conservación del patrimonio natural (pp. 11-21). Ministerio de Ciencia, Tecnología y Medio Ambiente (CITMA).
ONU (2025). La pérdida de biodiversidad exige medidas urgentes a escala mundial. En: https://news.un.org/es/story/2025/05/1538951.
Rosenberg, K. V., Dokter, A. M., Blancher, P. J., Sauer, J. R., Smith, A. C., Smith, P. A., Stanton, J. C., Panjabi, A., Helft, L., Parr, M., & Marra, P. P. (2019). Decline of the North American avifauna. Science, 366(6461), 120–124. https://doi.org/10.1126/science.aaw1313.